| Les reins des Vertébrés : une diversité fonctionnelle sur un même thème structural | ||
|---|---|---|
| Précédent | La diversité des reins des Vertébrés | Suivant |
Le fonctionnement du néphron des Vertébrés peut être décrit à partir de l’exemple des Mammifères. Ceux des néphrons des autres Vertébrés peuvent ensuite lui être comparées.
De manières générale, la production de l’urine comporte deux phases : la formation d’une urine qualifiée de primitive et sa transformation en urine dite définitive, émise dans le milieu.
Une diversité de production de l’urine primitive
Copyright : 2019 – Mélissa Degauque ; Laure Del Rosso ; Driss Fassih ; Tomy Sagnial ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.

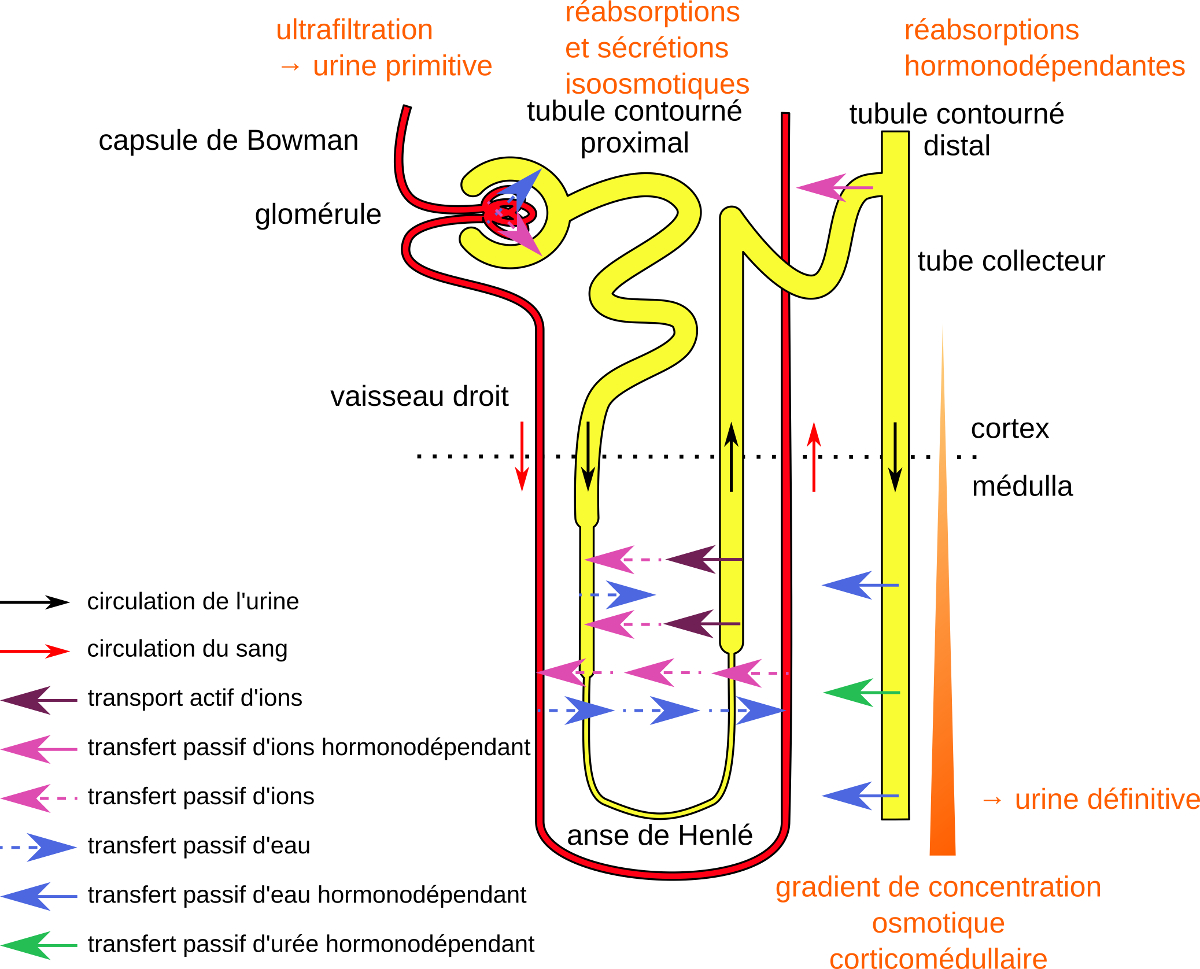
Chez les Mammifères, le corpuscule de Malpighi est une structure composite associant l’appareil circulatoire, avec le glomérule, et l’appareil excréteur, avec la capsule de Bowman du néphron.
Le glomérule est un bouquet de vaisseaux capillaires issus de la ramification d’une artériole afférente. Ils convergent en une artériole efférente irriguant les différentes régions du tubule urinaire, et formant en particulier des vaisseaux droits parallèles aux branches de l’anse de Henlé. Les vaisseaux capillaires issus de sa ramification convergent ensuite en veine rénale.
La capsule de Bowman comporte un feuillet externe, pariétal, et un feuillet interne, viscéral, enveloppant les vaisseaux capillaires du glomérule. L’espace de Bowman séparant les deux feuillets contient l’urine primitive.
Les pressions en présence au niveau du corpuscule de Malpighi sont :
- la pression sanguine, pression hydrostatique régnant dans les capillaires glomérulaires de l’ordre de 6,65 kPa ;
- la pression osmotique due notamment aux molécules protéiques présentes dans le plasma de l’ordre de 4,00 kPa ;
- la pression intracapsulaire, pression hydrostatique régnant dans l’espace de Bowman de l’ordre de 1,33 kPa.
La première tend à faire sortir le sang des vaisseaux capillaires alors que la deuxième et la troisième s’y opposent.
La pression résultante est de l’ordre de 1,32 kPa. Elle provoque la sortie du sang des vaisseaux capillaires en direction de l’espace de Bowman.
Le compartiment sanguin et le compatiment urinaire sont séparés par la paroi des vaisseaux capillaires, une membrane basale et le feuillet viscéral. L’ensemble constitue un filtre traversé par le liquide sanguin. Les dimensions de ses mailles sont telles que les cellules sanguines et les molécules de grande taille comme les protéines sont retenues. Seuls l’eau et les solutés de dimensions réduites, en particulier les déchets du métabolisme, passent des vaisseaux capillaires à l’espace de Bowman à travers cette barrière de filtration. Le liquide recueilli dans l’espace de Bowman est l’urine primitive.
L’urine primitive est donc produite par une ultrafiltration du sang, due à une différence de pression entre les compartiments sanguin et urinaire, à travers la barrière de filtration que constituent la paroi des vaisseaux capillaires, la membrane basale et le feuillet viscéral de la capsule de Bowman.
La production de l’urine primitive ainsi décrite est rencontrée dans tous les néphrons à glomérule interne, présents chez les Sélaciens, les Téléostéens, les Lissamphibiens, les Sauropsidés et les Mammifères.
Elle présente une relative diversité quant à la pression et au débit sanguins dans le glomérule, déterminant le débit de filtration glomérulaire (volume d’urine primitive formé par unité de temps). Il est ainsi important chez les Mammifères en raison d’une pression et d’un débit sanguin élevés, plus faible chez les Téléostéens en raison d’une pression plus basse.
En revanche, quelle que soit l’irrigation du rein, le glomérule est toujours issu de la ramification de l’artère rénale.
Dans le cas du néphron à glomérule externe décrit chez les larves de Lissamphibiens, il n’existe pas d’association étroite entre glomérule et néphron. Le glomérule est associé à la paroi de la cavité cœlomique.
Du fait d’une différence de pression entre le sang et le liquide cœlomique, le liquide sanguin tend à passer dans la cavité cœlomique. Le sang est filtré à travers la paroi des vaisseaux capillaires, une membrane basale et le feuillet délimitant la cavité cœlomique.
Les battements des cils du néphrostome mettent en mouvement le liquide cœlomique en direction du tubule urinaire. Il devient l’urine primitive, qui est donc formée par ultrafiltration et déplacement vers le néphron.
Le néphron aglomérulé est constitué du seul tubule urinaire, fermé à une extrémité et ouvert sur un tube collecteur à l’autre.
La production d’urine primitive implique un transport actif d’ions, le plus souvent divalents, du milieu intérieur vers la lumière du tubule. L’accumulation des ions dans la lumière du tubule est à l’origine d’un gradient de concentration osmotique provoquant l’entrée d’eau dans le tubule.
La formation de l’urine primitive n’implique pas de filtration, mais des mouvements osmotiques.
Comment l’urine primitive est-elle transformée en urine définitive ?
Une diversité de production de l’urine définitive
Les Mammifères éliminent une urine définitive dont la concentration osmotique est supérieure à celle de l’urine primitive et du plasma, et dont la composition est différente.
L’urine primitive étant produite par une ultrafiltration non sélective pour les solutés de petite taille, il est vraisemblable que des processus de réabsorptions et de sécrétions interviennent dans le tubule urinaire.
Le tubule contourné proximal est le siège d’une réabsorption active de multiples molécules organiques comme le glucose, les acides aminés, le lactate, ainsi que de substances minérales comme les ions Na+ et HCO3–.
La réabsorption est à l’origine d’un gradient de concentration osmotique entre le compartiment urinaire et le milieu intérieur, responsable d’une réabsorption importante d’eau.
Les concentrations osmotiques de l’urine à son entrée et à sa sortie du tubule contourné proximal étant similaires, les réabsorptions sont qualifiées d’isoosmotiques.
Le tubule contourné distal est également le siège d’une réabsorption d’ions Na+ mais elle est contrôlée par une hormone, l’aldostérone. Une sécrétion d’ions K+ intervient aussi.
Par ailleurs, des ions ammonium sont sécrétés sur toute la longueur du tubule urinaire.
L’anse de Henlé est formée d’un segment large descendant, dans la continuité du tubule contourné proximal, d’un segment grêle descendant, d’un segment grêle ascendant puis d’un segment large ascendant en continuité avec le tubule contourné distal.
Le segment large descendant est perméable aux ions Na+ et Cl– et / ou à l’eau, le segment grêle descendant est imperméable aux solutés mais perméable à l’eau, le segment grêle ascendant est perméable aux solutés mais imperméable à l’eau, tandis que le segment large ascendant réalise un transport actif d’ions Na+ et Cl– de l’urine au milieu intérieur tout en étant imperméable à l’eau.
Schématiquement, les ions Na+ et Cl– expulsés depuis la lumière du segment large ascendant s’accumulent dans l’interstitium et entrent passivement dans la lumière du segment descendant, ils subissent ainsi un recyclage. Les segments descendant et ascendant cheminant parallèlement, ils déterminent un dispositif à contre-courant. L’enrichissement de l’interstitum en ions Na+ et Cl– s’en trouve augmenté en direction de la pointe de l’anse de Henlé.
Un gradient de concentration osmotique corticomédullaire est ainsi établi.
Les tubes collecteurs acheminent l’urine dans le sens cortex-médulla en direction des uretères. Ils rencontrent un environnement dont la concentration osmotique croît progressivement. Or leur paroi est perméable à l’eau en présence de l’hormone anti-diurétique (ADH). En conséquence, l’eau est réabsorbée par les tubes collecteurs au fur et à mesure de leur parcours vers la médulla profonde et l’urine concentrée, jusqu’à devenir hyperosmotique par rapport au plasma.
Finalement, l’urine primitive est transformée en urine définitive par un ensemble de processus de réabsoptions et sécrétions, le plus souvent actives, réalisées par le tubule urinaire.
Le devenir de l’eau de l’urine primitive est très variable chez les Vertébrés.
L’anse de Henlé est responsable de l’établissement d’un gradient osmotique à l’origine de la concentration finale de l’urine par réabsorption d’eau.
Les néphrons qui en sont dépourvus ne peuvent produire une urine de concentration osmotique supérieure à celle du plasma. La plupart des Vertébrés sont concernés, exception faite des Mammifères et des Oiseaux.
Par ailleurs, les capacités de concentration de l’urine sont variées au sein d’un même groupe, ainsi les Mammifères déserticoles produisent une urine dont la concentration osmotique est supérieure à celle des Mammifères vivant dans un milieu riche en eau.
La concentration finale de l’urine contribue à l’équilibre hydrique de l’organisme, permettant une réduction des pertes.
Inversement, les Téléostéens d’eau douce éliminent une urine abondante et diluée.
Leurs néphrons réalisent une réabsorption réduite de l’eau de l’urine primitive, produite en grande quantité. Ils évacuent ainsi l’eau excédentaire entrée dans leur organisme par osmose. En revanche une réabsorption des ions, notamment monovalents, est effectuée par les tubules urinaires, proximaux en particulier.
Les Téléostéens marins quant à eux éliminent une urine peu abondante et peu diluée.
Les ions excédentaires, notamment divalents, sont sécrétés par les tubules proximaux des néphrons, souvent seuls présents.
Chez les Sélaciens marins, l’urine éliminée est relativement concentrée et moyennement abondante.
Les tubules proximaux des néphrons sécrètent des ions, divalents principalement, ainsi qu’un peu d’eau, et les tubules distaux réabsorbent l’urée composé essentiel du plasma.
Ainsi, la modification de la composition de l’urine primitive par les néphrons desVertébrés implique généralement des processus actifs, et varie avec les contraintes hydriques et ioniques que connaissent les espèces en relation avec leurs milieux de vie. Les néphrons contribuent à éliminer les déchets azotés mais aussi au maintien de l’équilibre hydrique et ionique.
Dans cette perspective, le fonctionnement des néphrons aglomérulés peut être interprété comme un moyen de contrôler la quantité et la composition de l’urine primitive, réduisant ainsi les réabsorptions et sécrétions coûteuses en énergie.