| De la circulation sanguine simple à la circulation sanguine double : une distribution du sang dans un circuit partiellement dédoublé | ||
|---|---|---|
| Précédent | Les circulations sanguines simple et double chez les Vertébrés | Suivant |
Parmi les Vertébrés, outre les groupes strictement aquatiques ou aériens, existent desgroupes occupant à la fois les milieux aquatique et aérien.
Ce sont par exemple les Lissamphibiens comme les Grenouilles, mais aussi certainsVertébrés aquatiques comme les Dipneustes capables de réaliser leurs échanges gazeuxrespiratoires avec l’eau par des branchies ou avec l’air par des poumons.
De la circulation sanguine simple à la circulation sanguine double : un circuit double et un cœur partiellement cloisonné
L’exemple de la Grenouille permet de décrire le cas des Lissamphibiens.
Copyright : 2019 Marc Second Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.

Copyright : 2019 Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
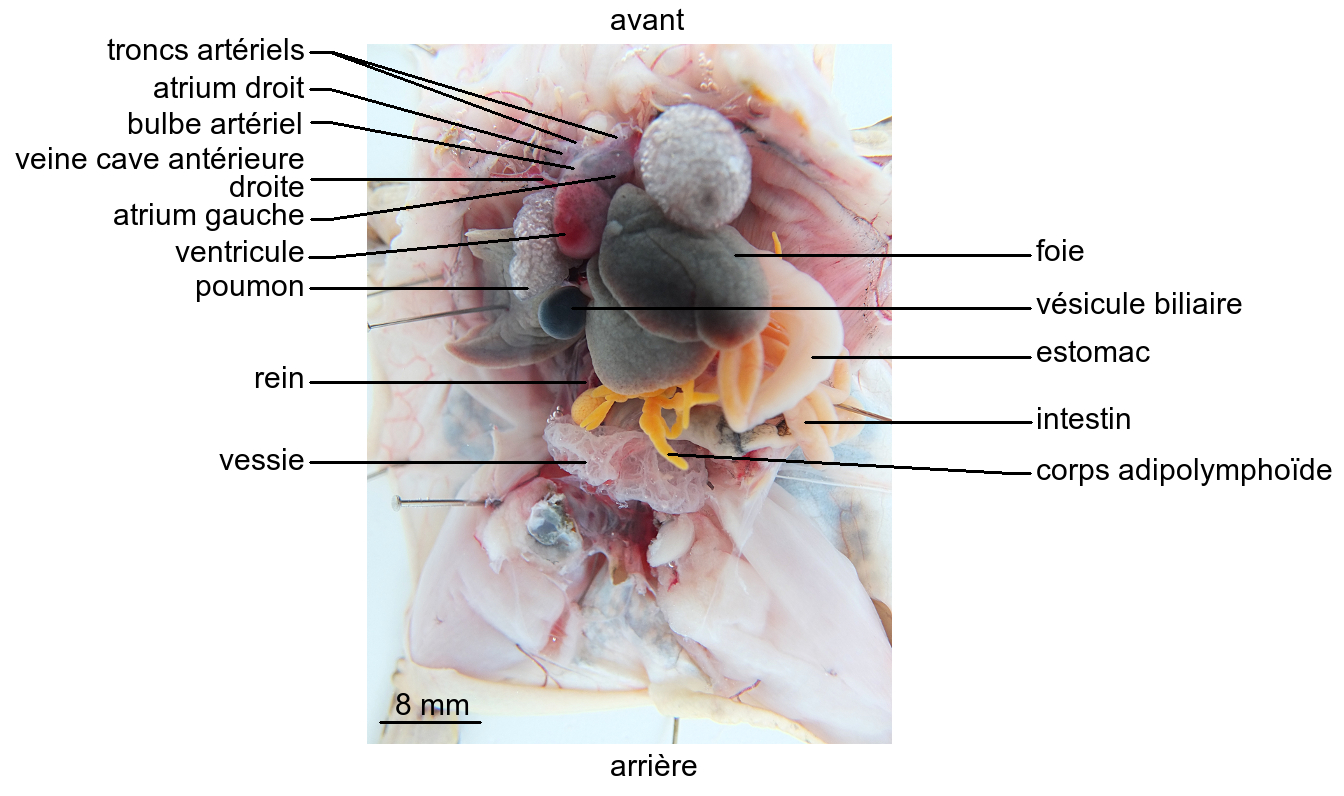
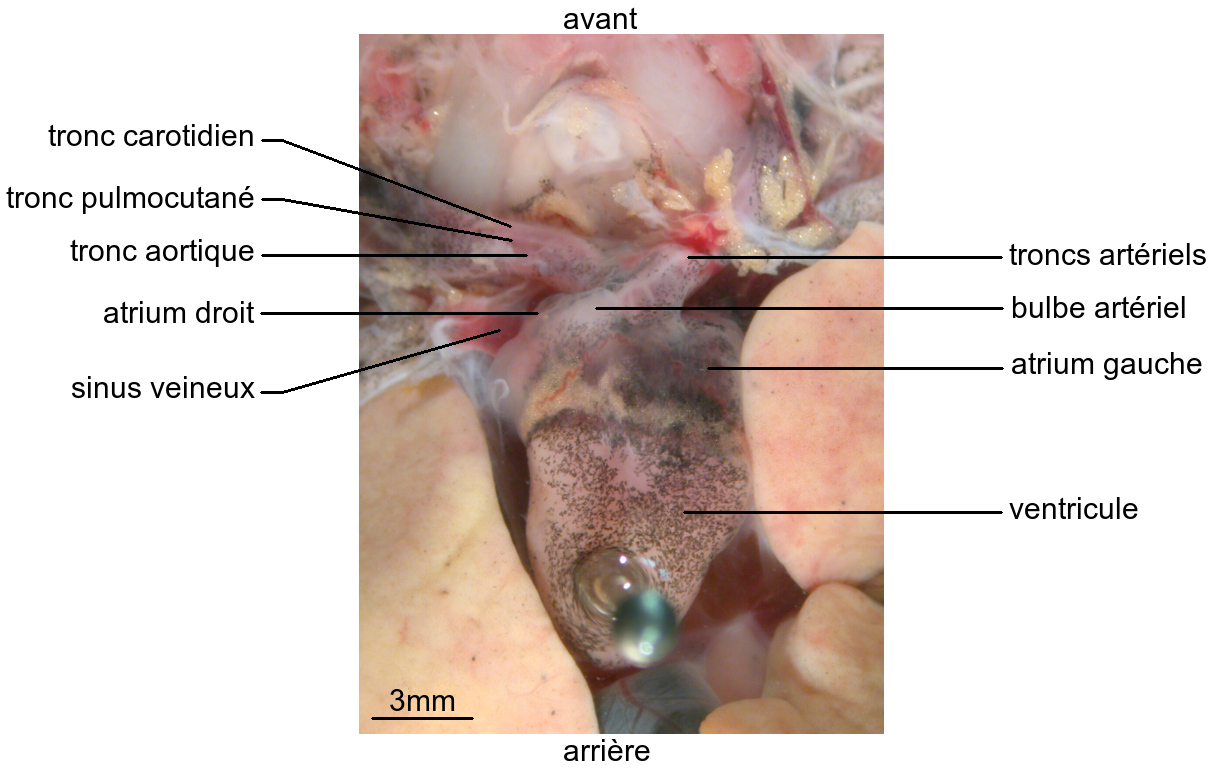
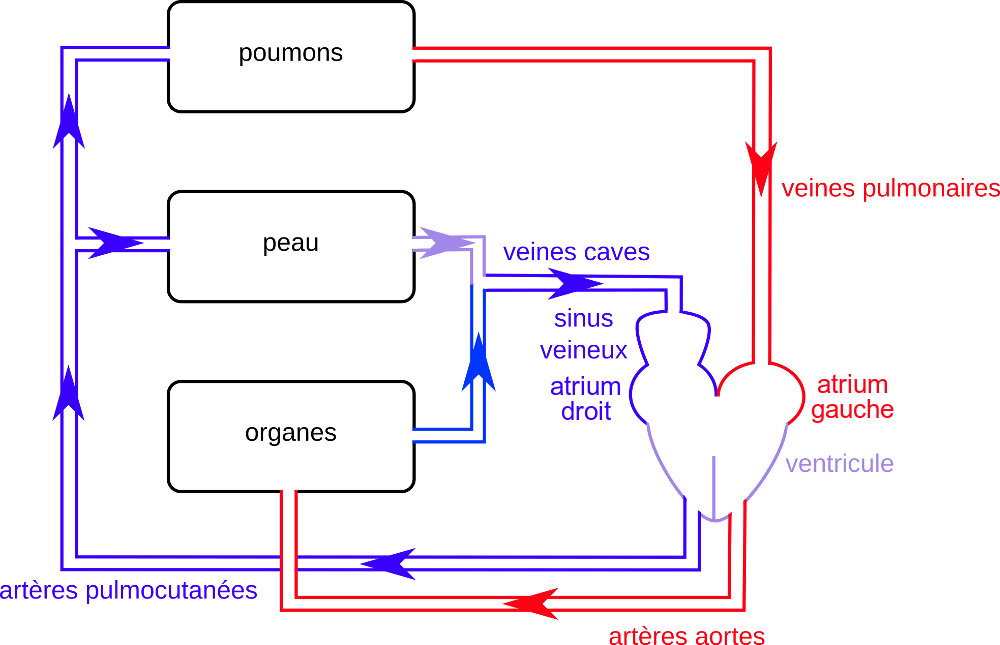
L’appareil circulatoire de la Grenouille adulte comporte un cœur situé en position antérieure et ventrale. Anatomiquement, il apparaît formé d’un atrium droit communiquant avec un sinus veineux dorsal, d’un atrium gauche et d’un ventricule. Un bulbe artériel ventral est en continuité avec le ventricule.
Le bulbe artériel reçoit le sang du ventricule. De chaque côté, il se ramifie en trois vaisseaux ou troncs artériels :
- un tronc carotidien se dirigeant vers l’avant et irriguant la tête ;
- un tronc aortique courbé vers l’arrière, formant une crosse aortique, et acheminant le sang vers les organes de la région postérieure ;
- un tronc pulmocutané latéral apportant le sang aux poumons d’une part et à la peau d’autre part.
Dorsalement, les deux artères aortes convergent en une artère aorte unique émettant une artère cœliacomésentérique en direction de l’appareil digestif, des artères rénales alimentant les reins et des artères iliaques irriguant les membres postérieurs.
Le sang des poumons est ramené à l’atrium gauche par deux veines pulmonaires, alors que le sang issu des autres organes est drainé par des veines caves antérieures et postérieure débouchant dans le sinus veineux.
Dans les poumons et la peau, le sang est enrichi en dioxygène et le dioxyde de carbone qu’il contient est libéré dans le milieu. Dans les autres organes, il est appauvri en dioxygène et enrichi en dioxyde de carbone.
Finalement, l’atrium gauche reçoit du sang hématosé et le sinus veineux un mélange de sang hématosé provenant de la peau et de sang non hématosé provenant des autres organes.
Copyright : 2019 Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
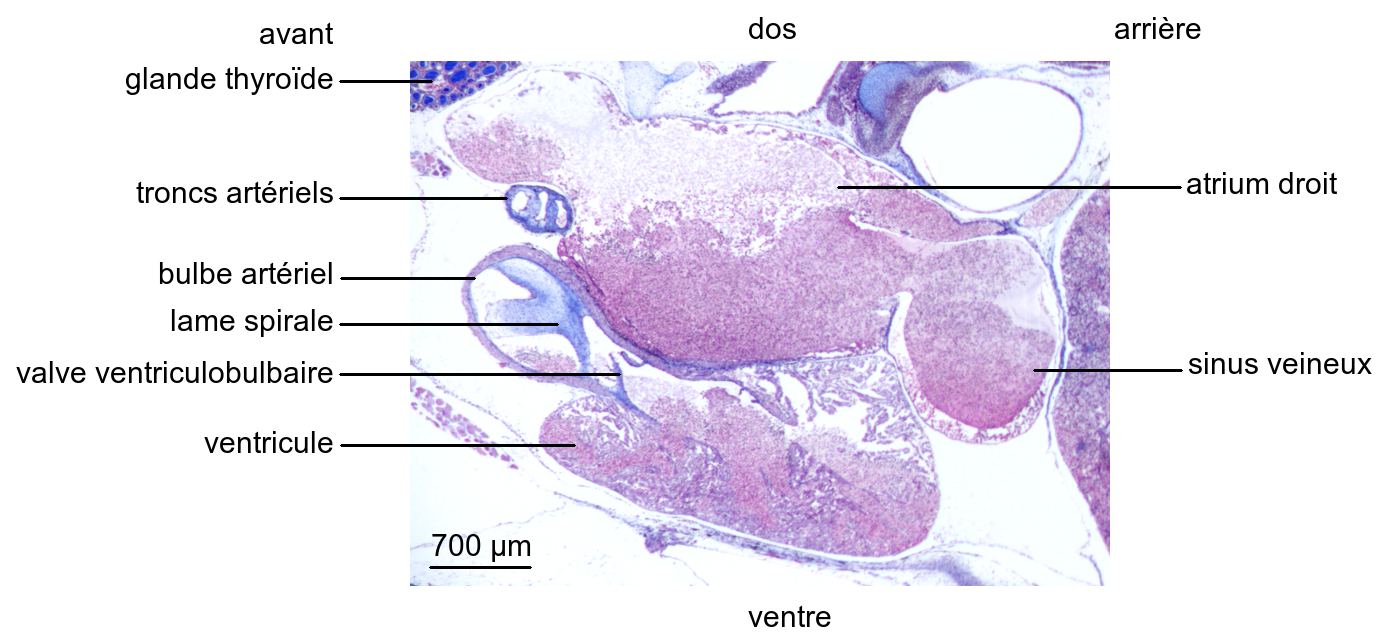
En coupe longitudinale, il apparaît que le sinus veineux est en continuité avec l’atrium droit et que les atriums droit et gauche communiquent avec le ventricule qui demeure indivis. Le bulbe artériel pour sa part est divisé en une rampe dorsale et une rampe ventrale par une lame spirale de nature conjonctive.
Ainsi le cœur de la Grenouille est incomplètement cloisonné, avec deux atriums distincts et un ventricule commun. Une telle organisation rend possible le mélange des sangs hématosé et non hématosés dans le ventricule. Cependant, le sang hématosé provenant des poumons et transitant par l’atrium gauche est principalement dirigé vers la rampe ventrale du bulbe artériel, à l’origine des troncs carotidien et aortiques.
Le sang résultant du mélange de sang hématosé cutané et de sang non hématosé systémique transite par le sinus veineux et l’atrium droit avant d’être propulsé dans la rampe dorsale du bulbe artériel, à l’origine du tronc pulmoncutané.
Il existe un cloisonnement fonctionnel du ventricule résultant notamment d’un léger décalage dans le remplissage des atriums gauche et droit, ainsi que d’une orientation des flux sanguins dans le ventricule du fait de la présence de replis dans sa paroi.
Avec une telle organisation, les réseaux capillaires de l’appareil respiratoire et des autres appareils sont disposés en parallèle.
Les débits sont contrôlés par le jeu des résistances vasculaires et il est possible de distribuer des volumes de sang différents dans les circuits pulmonaire et systémique.
Ainsi, en hiver alors qu’ils sont en vie ralentie, les Lissamphibiens réalisent leurs échanges gazeux respiratoires exclusivement par la peau. L’irrigation pulmonaire est alors réduite par une augmentation de la résistance vasculaire et le sang principalement orienté vers la peau et les autres appareils. Le sang hématosé au niveau de la peau parvient à l’atrium droit, est mélangé au sang non hématosé puis redistribué entre la peau et la circulation systémique.
La circulation des Lissamphibiens peut être représentée comme ci-dessous.
Copyright : 2019 Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
L’étude du développement de l’appareil circulatoire des Lissamphibiens révèle que les troncs pulmocutané, aortique et carotidien sont homologues des artères branchiales 6, 4 et 3 des Téléostéens, les réseaux capillaires présents sur leurs trajets ayant disparu.
De la même manière chez les Mammifères et les Oiseaux, les artères pulmonaires sont une évolution des artères branchiales 6, l’artère aorte de l’une des crosses aortiques issue des artères branchiales 4 (gauche chez les Mammifères et droite chez les Oiseaux) et les artères carotides des artères branchiales 3.
Il est vraisemblable qu’il existe des états intermédiaires entre ceux observés chez les Téléostéens et les Lissamhibiens d’une part, entre les Lissamphibiens et les Mammifères et Oiseaux d’autre part. Quelles sont leurs caractéristiques ?
De la circulation sanguine simple à la circulation sanguine double : vers un circuit dédoublé et un cœur cloisonné
Les Téléostéens appartiennent au sous-groupe de Vertébrés des Actinoptérygiens.
Le sous-groupe des Sarcoptérygiens est principalement représenté par des animaux
évoluant en milieu aérien dont les Mammifères, les Oiseaux et les Lissamphibiens. Il
comporte également des animaux aquatiques, les Dipneustes, qui ont la particularité
de posséder un double appareil respiratoire, branchial et pulmonaire.
Copyright : 2019 Marc Second
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
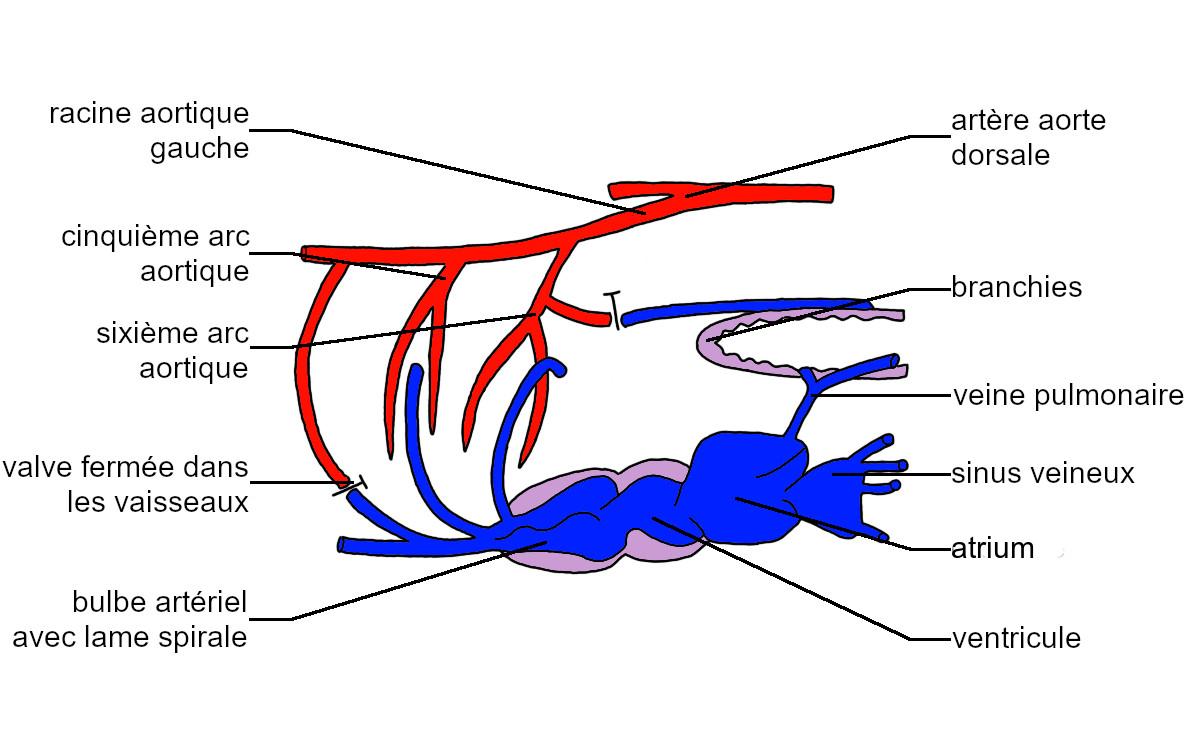
À l’instar de celui des Téléostéens, l’appareil circulatoire des Dipneustes est représenté par cinq paires d’artères antérieures, correspondant aux artères branchiales embryonnaires 2 à 5.
Seules les artères 2, 5 et 6 sont associées à des branchies, les artères 3 et 4 sont continues. La sixième artère branchiale efférente émet une ramification en direction des poumons, qualifiée d’artère pulmonaire. Le sang du poumon retourne au cœur par une veine pulmonaire.
Parallèlement le cœur, formé de quatre chambres semblables à celles du cœur des Téléostéens, comporte une cloison atriale et une cloison ventriculaire incomplètes. Elles déterminent des régions cardiaques droite et gauche. Le bulbe artériel est cloisonné de même par une lame spirale.
Lorsque le milieu aquatique est riche en dioxygène, les échanges gazeux respiratoires et la circulation sont semblables à ceux décrits chez les Téléostéens : le sang pauvre en dioxygène est propulsé dans les artères branchiales afférentes, il est hématosé dans les branchies, repris par les artères branchiales efférentes puis distribué aux autres organes. Appauvri en dioxygène, il retourne au cœur.
Dans cette situation, le passage du sang dans les poumons est empêché par la fermeture de la valve située au niveau des artères pulmonaires.
Lorsque le milieu aquatique est pauvre en dioxygène, les Dipneustes réalisent leurs échanges gazeux respiratoires avec l’air grâce à leurs poumons. Les valves des artères branchiales 2 et 3 sont fermées et le sang propulsé par le cœur est orienté vers l’artère branchiale afférente 6 puis les poumons. Le sang hématosé est alors ramené au sinus veineux par les veines pulmonaires. Il circule dans la moitié gauche du cœur et est propulsé dans les artères 3 et 4 qui le distribuent vers la tête et la région postérieure. Le sang non hématosé retourne au cœur par des veines systémiques et circule dans la région droite avant d’être à nouveau propulsé dans l’artère 6 et pris en charge par l’artère pulmonaire.
La circulation ainsi décrite est modulable : selon les conditions environnementales, elle est simple ou double. Deux circuits coexistent grâce à la présence de valves vasculaires et de cloisons cardiaques incomplètes.
La circulation simple rappelle celle des Vertébrés aquatiques et la circulation double celle des Lissamphibiens, suggérant une préadaptation des Dipneustes à la vie en milieu aérien.
La coexistence des deux types de circulation révèle les relations étroites entre l’organisation de l’appareil circulatoire et celle de l’appareil respiratoire.
Parmi les Vertébrés évoluant en milieu aérien figurent les Crocodiliens comme les Crocodiles, les Chéloniens ou Tortues et les Lépidosauriens avec les Lézards par exemple.
L’appareil circulatoire des Chéloniens et des Lépidosauriens est semblable à celui des Lissamphibiens, exception faite de la dérivation cutanée de l’artère pulmonaire. La peau est dans ces groupes irriguée par le circuit systémique. Le sinus veineux est réduit et le ventricule partiellement cloisonné.
Un tel agencement permet de distribuer le sang différemment selon les conditions, notamment chez les Chéloniens évoluant en milieu aquatique.
Ainsi, lorsque l’animal est en surface et que les poumons sont ventilés, la résistance vasculaire pulmonaire est réduite et le volume de sang dans la circulation pulmonaire plus élevé que dans la circulation systémique. Le sang est alors très enrichi en dioxygène, étant partiellement recyclé dans la circulation pulmonaire.
Inversement en plongée, lorsque les poumons ne sont pas ventilés, la résistance vasculaire pulmonaire augmente et le volume de sang dans la circulation pulmonaire est plus faible que dans la circulation systémique. Un mélange de sang non hématosé et de sang hématosé intervient, le sang étant partiellement recyclé au sein de la circulation systémique. Ce fonctionnement assure un approvisionnement stable des organes en dioxygène.
Copyright : 2019 Marc Second
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
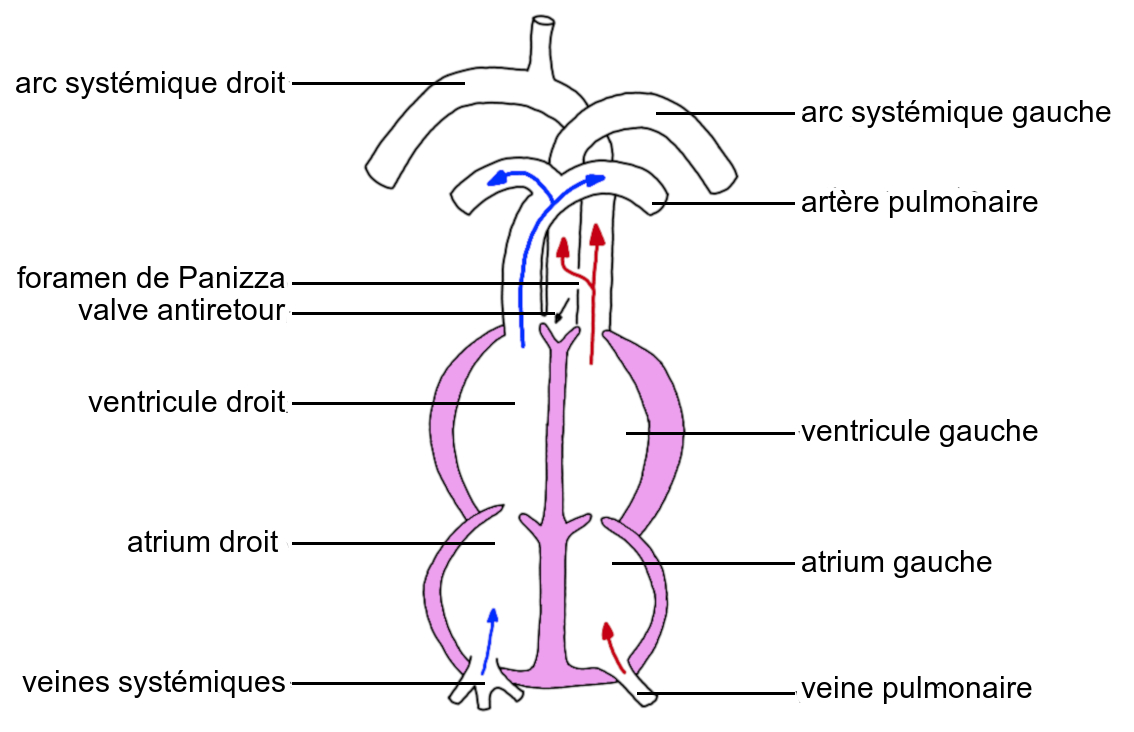
Le cœur des Crocodiliens présente en revanche une cloison interventriculaire continue, comme ceux des Mammifères et des Oiseaux. Cependant, si l’artère systémique droite communique avec le ventricule gauche, l’artère systémique gauche communique avec le ventricule droit. Les deux artères systémiques sont reliées par un conduit appelé foramen de Panizza.
Lors de la systole des ventricules, le ventricule gauche propulse le sang hématosé dans l’artère systémique droite, et du fait de la pression, une partie du sang hématosé passe dans l’artère systémique gauche en empruntant le foramen de Panizza. Parallèlement, la valve située à la base de l’artère systémique gauche est fermée, et le ventricule droit propulse le sang non hématosé dans les seules artères pulmonaires.
Une double circulation est ainsi établie, avec la séparation anatomique complète des circuits pulmonaire et systémique.