| Les échanges gazeux respiratoires des Euarthropodes en milieu aquatique | ||
|---|---|---|
| Précédent | Les échanges gazeux respiratoires des Euarthropodes | Suivant |
Parmi les Euarthropodes aquatiques figurent les Malacostracés dont le Crabe vert et l’Écrevisse sont des exemples.
Les organes et les modalités des échanges de gaz respiratoires en milieu aquatique peuvent être décrits pour ces espèces.
Les branchies, des expansions tégumentaires latérales réalisant les échanges gazeux respiratoires

Copyright : 2019 – Émilie Bresson ; Élysa Chavarin ; Charlotte Décline Emonot ; Céline Dugat ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.

Le Crabe vert possède deux cavités branchiales latérales, situées de part et d’autre du céphalothorax à l’extérieur des parois corporelles latérales, les pleurites. Elles sont délimitées par des expansions latérales du tégument dorsal, les branchiostégites. Ventralement, les branchiostégites sont soudés au tégument mais des orifices sont ménagés à la base des pattes antérieures.
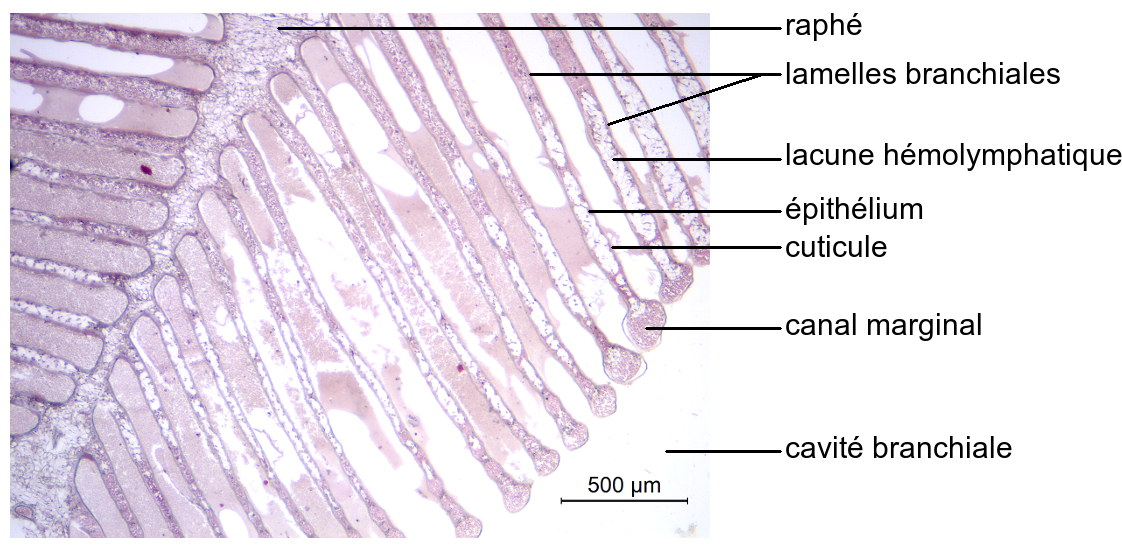
Dans chaque cavité branchiale neuf branchies sont présentes, insérées ventralement sur le pleurite, les membranes articulaires ou les pattes. Elles sont respectivement appelées pleurobranchies, arthrobranchies et podobranchies. Leur extrémité libre, dorsale, est effilée.
Les branchies comportent un axe portant deux rangées de lamelles latérales.
Copyright : 2019 – Émilie Bresson ; Élysa Chavarin ; Charlotte Décline Emonot ; Céline Dugat ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
Sur l’axe de la branchie, appelé raphé, de multiples lamelles plates sont insérées de chaque côté.
Parallèles et peu épaisses, elles sont délimitées par une fine paroi constituée d’un épithélium simple et cubique surmonté d’une cuticule. Il s’agit d’expansions repliées du tégument, dont la cuticule est devenue très fine. Leur extrémité est légèrement renflée et abrite un canal marginal. L’espace situé entre les deux épithéliums contient de l’hémolymphe, liquide extracellulaire circulant.
En relation avec la présence de lamelles plates portées par un axe, les branchies du Crabe vert sont appelées phyllobranchies.
Copyright : 2019 – Émilie Bresson ; Élysa Chavarin ; Charlotte Décline Emonot ; Céline Dugat ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
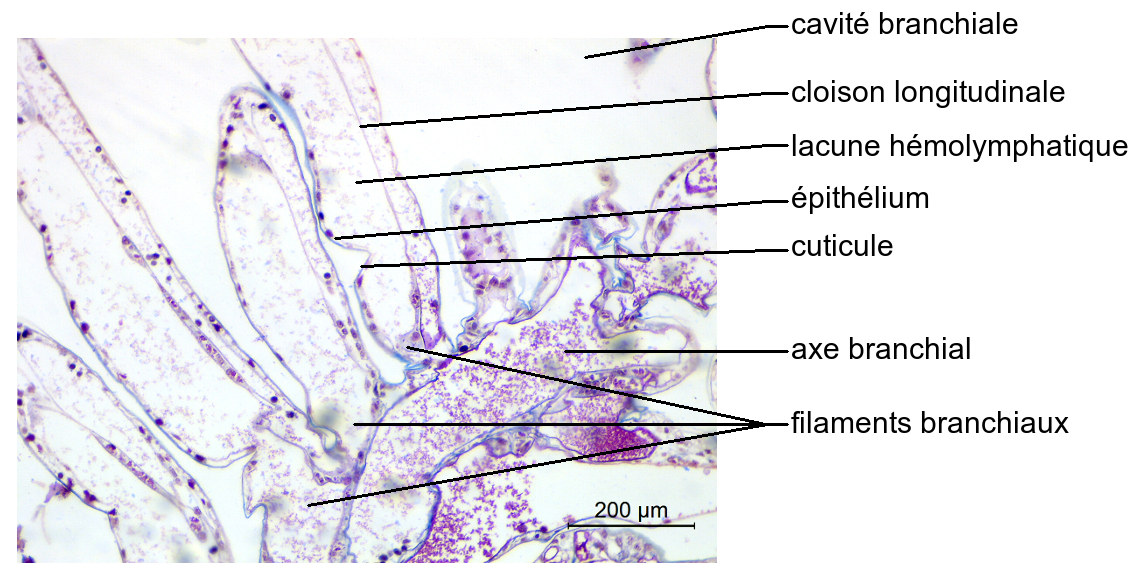
De même que le Crabe vert, l’Écrevisse possède des branchies latérales logées dans des cavités branchiales protégées par des branchiostégites.
Elles sont formées de filaments portés par un axe. Leur paroi est fine, constituée d’un épithélium simple et cubique surmonté d’une cuticule peu épaisse. Elles contiennent une lacune dans laquelle circule l’hémolymphe, séparée en deux compartiments par une cloison longitudinale.
En raison de la présence de filaments insérés sur un axe, les branchies de l’Écrevisse sont appelées trichobranchies.
Finalement les organes respiratoires des Malacostracés, et plus généralement des Euarthropodes aquatiques, sont des branchies, expansions tégumentaires de faible épaisseur en forme de lamelles ou de filaments.
En raison de leur nombre élevé, la surface de contact des branchies avec l’eau contenue dans la cavité branchiale est grande.
Ces expansions sont soutenues par l’eau environnante, à l’origine d’une poussée d’Archimède importante liée à sa densité.
Fragiles du fait de leur faible épaisseur, elles sont par ailleurs protégées par les expansions du tégument délimitant les cavités branchiales.
Au cœur des lamelles et des filaments branchiaux des lacunes contenant de l’hémolymphe, liquide extracellulaire circulant des Euarthropodes, sont présentes.
L’échangeur des gaz respiratoires est représenté par l’épithélium et la cuticule séparant le milieu extérieur du milieu intérieur. Son épaisseur est de l’ordre de 10µm. Il est traversé par le dioxygène, dans le sens milieu vers hémolymphe et par le dioxyde de carbone en sens inverse.
La ventilation et la circulation, renouvellements des milieux extérieur et intérieur des branchies
Écouter/Voir la vidéo
au format .mp4
au format .webm
Copyright 2019 – Émilie Bresson ; Élysa Chavarin ; Charlotte Décline Emonot ; Céline Dugat
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
Le dépôt de bleu de méthylène au niveau des orifices des cavités branchiales du Crabe vert, situés à la base des pattes antérieures ventralement, conduit à une expulsion de bleu de méthylène par le cadre buccal.
Le flux de bleu de méthylène révèle l’existence d’une circulation d’eau dans la cavité branchiale, avec une entrée par les orifices ventraux et une sortie par le cadre buccal. Elle permet le renouvellement de l’eau de la cavité branchiale et porte le nom de ventilation.
La ventilation renouvelle l’eau baignant les branchies.
L’eau entrant apporte du dioxygène dissous alors que l’eau de la cavité branchiale est appauvrie en dioxygène du fait de son absorption.
L’eau sortant est inversement enrichie en dioxyde de carbone rejeté par l’organisme, qui se trouve ainsi évacué.
La ventilation est due aux battements des scaphognathites dans les cavités branchiales. Ce sont des expansions lamelleuses d’appendices buccaux, les maxilles.
Dans les cavités branchiales, l’eau effectue un circuit simple. La viscosité de l’eau étant importante, le coût énergétique de sa mise en mouvement est élevé. Dans ces conditions, une circulation selon un sens unique est plus économe en énergie qu’une circulation en deux sens. La ventilation est qualifiée d’unidirectionnelle.
Le Crabe vert vit dans la zone intertidale, soumise aux marées.
Lorsqu’il est émergé, il conserve de l’eau dans ses cavités branchiales, notamment entre les lamelles, évitant ainsi leur collapsus et la dessiccation. La ventilation est maintenue, mais les battements des scaphognathites sont inversés : leurs mouvements font barboter l’air du cadre buccal dans l’eau des cavités branchiales, assurant ainsi le renouvellement du dioxygène.
L’eau du milieu extérieur circule dans les cavités branchiales autour des lamelles. Parallèlement, l’hémolymphe circule dans les lamelles. Elle prend en charge le dioxygène et le distribue dans l’organisme d’une part, et apporte le dioxyde de carbone produit par les cellules d’autre part.
Dans les branchies du Crabe vert, la circulation de l’hémolymphe dans les lamelles est réalisée parallèlement à celle de l’eau dans la cavité branchiale, selon un dispositif dit à contre-courant. L’hémolymphe arrivant dans la lamelle contient peu de dioxygène et rencontre une eau moyennement oxygénée en raison du transfert de dioxygène vers l’hémolymphe effectué en amont. Un transfert de dioxygène est tout de même possible de l’eau à l’hémolymphe. En aval, l’hémolymphe plus riche en dioxygène rencontre une eau plus oxygénée, le transfert de dioxygène est encore possible. Un tel dispositif autorise des échanges gazeux sur toute la longueur de l’échangeur et accroît ainsi l’extraction du dioxygène du milieu extérieur vers le milieu intérieur par rapport à un dispositif à co-courant.
Quel est le mécanisme physique impliqué dans les échanges branchiaux de gaz respiratoires ?
La diffusion, mécanisme des échanges gazeux respiratoires
Les échanges gazeux respiratoires à l’échelle de l’organisme sont réalisés par un échangeur situé à l’interface entre le milieu extérieur et le milieu intérieur.
Il est traversé par le dioxygène du milieu extérieur vers le milieu intérieur qui le distribue, et en sens inverse par le dioxyde de carbone.
Il n’existe pas de transporteurs de gaz dans les membranes biologiques.
Les gaz traversent les membranes et les cellules par un mécanisme de diffusion simple, dans le sens des gradients de pressions partielles, sans consommation d’énergie.
La diffusion est décrite par la loi de Fick. Pour les gaz respiratoires, elle est exprimée comme suit :
Jx = Kx . ΔPx . S / E
avec :
Jx : flux du gaz x en mol.s-1 ;
Kx : constante de diffusion du gaz x en mol.s-1.m-1.Pa-1 ;
ΔPx : différence de pression partielle du gaz x de part et d’autre de l’interface en Pa ;
S : aire de la surface d’échanges en m2 ;
E : épaisseur de la surface d’échanges en m.
Il apparaît que le flux de gaz est d’autant plus important que la surface de l’échangeur est grande et que son épaisseur est faible.
Les branchies des Malacostracés, avec leurs multiples lamelles ou filaments, dont la paroi est réduite à une fine cuticule et un épithélium simple et cubique, ont une structure favorable à la diffusion des gaz respiratoires.
Le moteur de la diffusion est la différence de pression partielle du gaz entre les milieux extérieur et intérieur. La ventilation, renouvellement du milieu extérieur dans la cavité branchiale, et la circulation, déplacement du milieu intérieur sous l’échangeur, contribuent à maintenir le gradient de pression partielle.
Par exemple pour le dioxygène, la première est responsable de l’apport au niveau de l’échangeur et la seconde de l’évacuation.
La circulation à contre-courant des milieux extérieur et intérieur de part et d’autre de l’échangeur a pour conséquence le maintien d’un gradient de pression partielle faible mais constant sur toute la longueur de l’échangeur, autorisant les échanges sur tout cette longueur.
Par comparaison, comment sont réalisés les échanges de gaz respiratoires chez les Euarthropodes aériens ?
Quelles sont les conséquences des contraintes du milieu aérien sur ces échanges, en relation avec la loi de Fick ?