| Les branchies : sites de flux spontanés et de transferts contrôlés d’eau et d’ions | ||
|---|---|---|
| Précédent | Le problème de l’eau et des ions chez les Téléostéens | Suivant |
Une surface importante et une faible épaisseur pour des flux d’ions spontanés
Les Vertébrés réalisent des échanges de matière et d’énergie avec leur milieu de vie. En particulier, ils y prélèvent du dioxygène et y rejettent du dioxyde de carbone. Le dioxygène est consommé et le dioxyde de carbone produit par une voie métabolique essentielle des cellules animales, la respiration cellulaire.
Ces échanges de gaz respiratoires entre l’organisme et le milieu se déroulent au niveau d’un appareil spécialisé, l’appareil respiratoire.
Le Vairon est un Téléostéen d’eau douce dont les branchies sont représentatives des organes respiratoires du groupe.
Copyright : 2019 – Aïmen Bouzidi ; Ronan Dadole ; Romain Fillâtre ; Thomas Lelièvre ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.

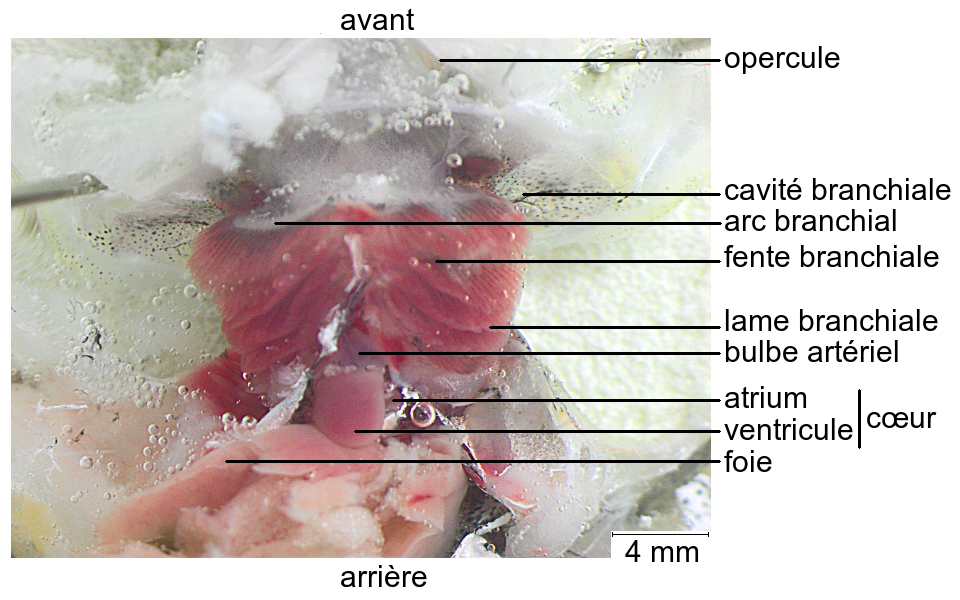
Les branchies sont des organes pairs, situés de chaque côté du pharynx à l’arrière du crâne, dans des cavités branchiales.
Chaque cavité branchiale est délimitée par une expansion du tégument, l’opercule, et contient quatre arcs branchiaux.
Deux rangées de lames branchiales, antérieure et postérieure, sont insérées sur chaque arc branchial. Les lames de la rangée postérieure d’un arc et celles de la rangée antérieure de l’arc suivant sont intercalées.
Copyright : 2019 – Aïmen Bouzidi ; Ronan Dadole ; Romain Fillâtre ; Thomas Lelièvre ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
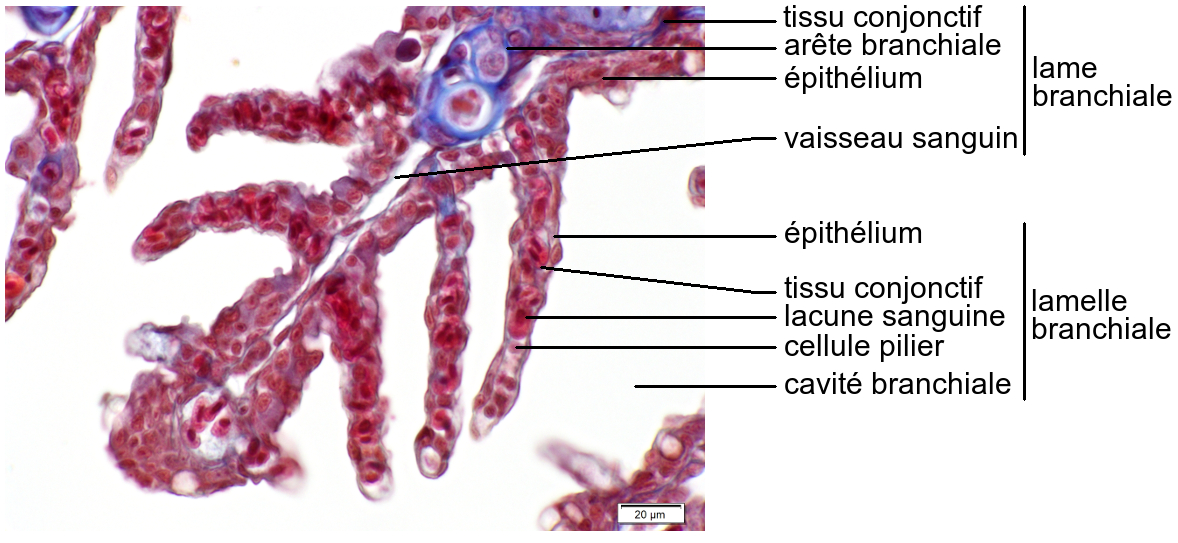
Les lames portent sur leurs deux faces de nombreuses lamelles branchiales, disposées perpendiculairement à leur surface.
Les lamelles sont constituées d’un épithélium simple pavimenteux replié sur lui-même, entourant de nombreuses lacunes sanguines soutenues par des cellules piliers. L’épithélium des lamelles branchiales est très fin, son épaisseur est de 2 µm environ. Les lamelles sont les sites des échanges gazeux respiratoires.
Les échanges gazeux respiratoires correspondent à une absorption de dioxygène et un rejet de dioxyde de carbone. Le dioxygène dissous dans le milieu extérieur rejoint le sang par diffusion simple à travers les cellules épithéliales. Le dioxyde de carbone fait le chemin inverse.
La diffusion simple est un transfert passif. La substance considérée diffuse d’un compartiment où elle est abondante vers un compartiment où elle est rare, sans dépense énergétique et sans action d’un transporteur spécialisé.
La loi de Fick décrit la diffusion simple. Pour un soluté x, elle est exprimée par la formule :
Jx = Dx x ΔCx x S / E
avec
- Jx, flux de diffusion du soluté en mol.s-1 ;
- Dx, coefficient de diffusion du soluté en m2.s-1 ;
- ΔPx, différence de concentration du soluté entre milieu extérieur et milieu intérieur en mol.m-3 ;
- S, aire de la surface d’échanges en m 2 ;
- E, épaisseur de la surface d’échanges en m.
Le flux de gaz est proportionnel à la différence de pression partielle entre les milieux extérieur et intérieur. Il est inversement proportionnel à l’épaisseur de l’échangeur, correspondant ici à l’épithélium des lamelles branchiales et au tissu conjonctif sous-jacent, et proportionnel à sa surface.
Présentant une épaisseur réduite et une surface importante, les branchies sont donc très favorables aux échanges passifs, de gaz respiratoires mais aussi d’autres substances.
Le milieu intérieur des Téléostéens vivant en eau douce est hyperosmotique et hyperionique par rapport à l’environnement et notamment à l’eau contenue dans les cavités branchiales.
Un flux spontané d’eau entrant et des flux spontanés d’ions sortants se produisent à travers l’échangeur des gaz respiratoires.
À l’inverse, le milieu intérieur des Téléostéens marins est hypoosmotique et hypoionique par rapport à l’eau de mer.
L’échangeur de gaz respiratoires branchial est dans ces conditions un site privilégié pour les flux spontanés sortant d’eau et entrants d’ions.
Comment les Téléostéens évoluant en eau douce compensent-ils le gain d’eau et les pertes d’ions ?
Comment les Téléostéens vivant en milieu marin compensent-ils la perte d’eau et les gains d’ions ?
Des ionocytes, des canaux et des pompes pour des transferts contrôlés d’eau et d’ions
Les transferts d’eau branchiaux sont réalisés à travers des canaux membranaires formés de protéines appelés aquaporines, dans le sens du gradient de concentration osmotique.
Dès lors, un contrôle partiel peut être exercé par l’animal sur le flux d’eau, notamment par le biais d’une modulation de l’expression des aquaporines.
Elle est stimulée par le cortisol, hormone produite par la glande interrénale, et en conséquence le flux d’eau branchial augmente. Inversement, elle est inhibée par la prolactine ce qui provoque une diminution du flux d’eau branchial. L’arginine-vasotocine imperméabilise également les branchies vis-à-vis de l’eau chez les Téléostéens d’eau douce.
Qu’en est-il des flux d’ions ?
Copyright : 2019 – Aïmen Bouzidi ; Ronan Dadole ; Romain Fillâtre ; Thomas Lelièvre ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
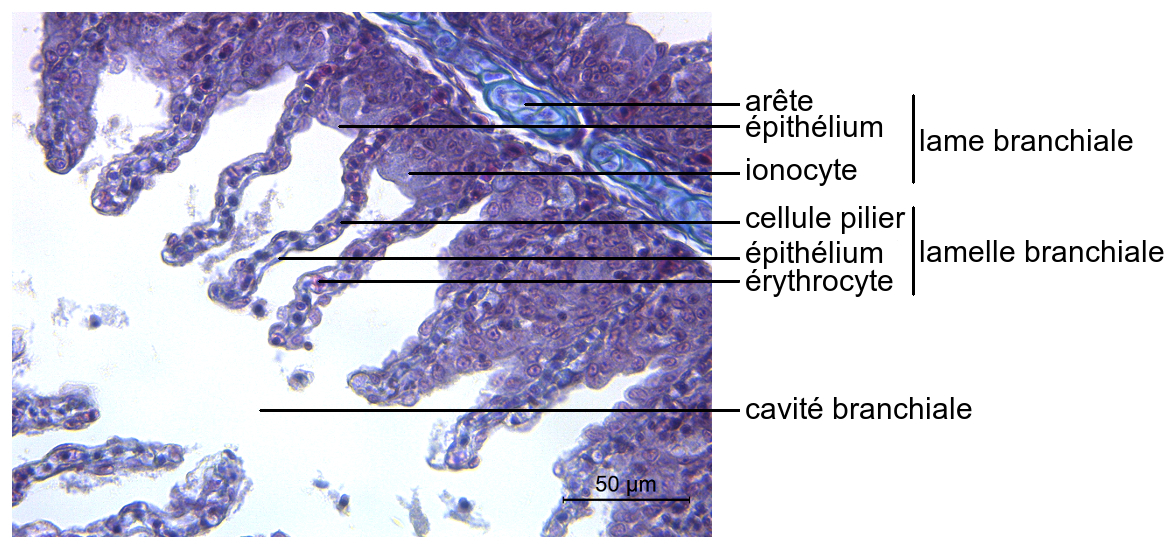
L’épithélium des lames branchiales des Téléostéens est principalement formé de cellules de revêtement, parmi lesquelles des cellules volumineuses, à cytoplasme clair et granuleux, sont dispersées.
Elles possèdent de nombreuses invaginations membranaires apicales et contiennent de multiples mitochondries. Divers canaux, transporteurs passifs et pompes ioniques sont présentes dans leur membrane, grâce auxquels elles réalisent des échanges d’ions.
Elles sont pour cette raison appelées ionocytes.
Copyright : 2019 – Ronan Dadole
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
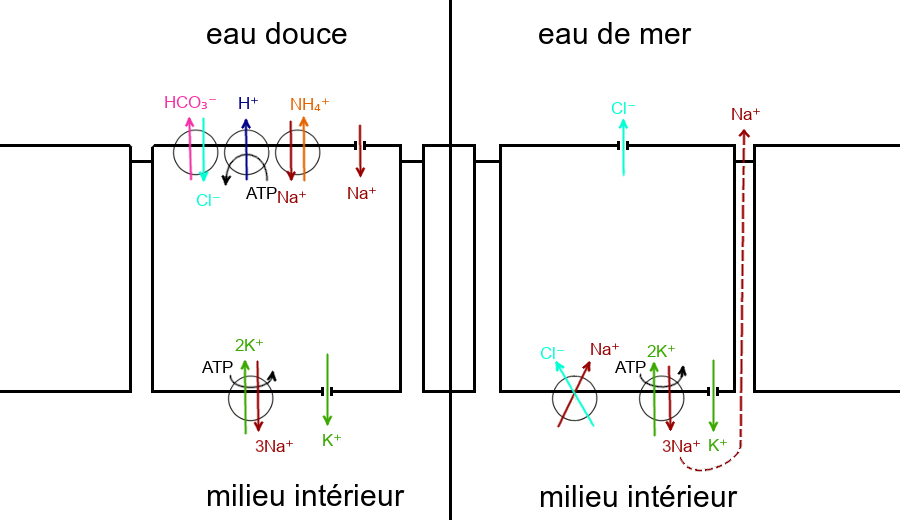
En eau douce, les cellules branchiales absorbent des ions Cl– et Na+ contre leurs gradients électrochimiques.
Les ions Cl– pénètrent dans la cellule alors que les ions HCO3 – en sortent, ils sont échangés grâce à un cotransporteur antiport apical.
Parallèlement, les ions Na+ pénètrent dans la cellule alors que les ions ammonium NH4 + en sortent, ils sont échangés grâce à un cotransporteur antiport apical.
Par ailleurs, une pompe ATPase Na+/K+ située dans la membrane plasmique du côté basal de la cellule expulse les ions Na+ hors de la cellule et fait entrer des ions K+, contre leurs gradients électrochimiques grâce à l’énergie fournie par l’hydrolyse de l’ATP.
Elle est à l’origine d’un gradient électrochimique transépithélial des ions Na+, provoquant leur entrée depuis le milieu extérieur à la faveur d’un canal apical, les ions K+ retournant au milieu intérieur grâce à un canal basal.
Un pompe ATPase H+ apicale expulsant les ions H+ pourrait contribuer à l’entrée passive des ions Na+ par l’établissement d’un gradient électrique la favorisant.
L’absorption des ions Cl– et Na+ contre les gradients électrochimiques par les cellules branchiales en eau douce implique une dépense d’énerge, nécessaire au fonctionnement des pompes ATPases.
Les cellules responsables des échanges décrits ne sont pas clairement identifiées chez les Téléostéens d’eau douce. Des flux contrôlés d’ions monovalents existant au niveau des lamelles branchiales, il est vraisemblable que l’épithélium des lamelles soit impliqué.
Les ionocytes des lames branchiales, appelés ionocytes β, interviendraient dans les flux des ions divalents en particulier des ions Ca2+.
En eau de mer, les ionocytes éliminent des ions Cl– et Na+ contre leurs gradients électrochimiques.
Une pompe ATPase Na+/K+ située dans la membrane plasmique du côté basal de la cellule expulse les ions Na+ hors de la cellule et fait entrer des ions K+.
Elle est à l’origine d’un gradient électrochimique transmembranaire des ions Na+, favorisant le fonctionnement d’un cotransporteur symport Na+/Cl– situé dans la membrane plasmique du côté basal.
Les ions K+ regagnent le milieu intérieur grâce à un canal basal.
Les ions Cl– gagnent le milieu extérieur à la faveur d’un canal membranaire situé à l’apex de la cellule, le CFTR (cystic fibrosis transmembrane regulator). Leur accumulation dans le milieu extérieur est à l’origine d’un gradient électrique provoquant le transfert des ions Na+ présents dans les espace intercellulaires vers le milieu extérieur.
De même qu’en eau douce, en milieu marin, la sécrétion des ions Cl– et Na+ contre leurs gradients électrochimiques implique une consommation d’énergie, permettant le fonctionnement de la pompe ATPase.
Les cellules responsables de la sécrétion sont les ionocytes des lames branchiales, appelés ionocytes α.
Ainsi, les branchies sont des organes effecteurs de l’osmorégulation, limitant les flux d’eau grâce à une imperméabilisation, et de l’ionorégulation, assurant l’absorption d’ions monovalents en eau douce et leur sécrétion en milieu marin.
Bien que les flux d’eau branchiaux sont limités, les Téléostéens marins perdent de l’eau et les Téléostens d’eau douce en gagnent.
Comment la perte d’eau est-elle compensée ?