| Les appareils branchiaux des Vertébrés : diversité et unité des convections des milieux intérieur et extérieur | ||
|---|---|---|
| Précédent | La diversité et l’unité des appareils branchiaux des Vertébrés | Suivant |
Une irrigation sanguine
Les lamelles et les filaments branchiaux des Vertébrés contiennent des lacunes ou des vaisseaux capillaires dans lesquels circule le sang, ils sont richement vascularisés.
Le dioxygène du milieu extérieur traverse l’échangeur en direction du sang, en raison du gradient de pression partielle. Le dioxyde de carbone suit le chemin inverse.
Ces diffusions ont pour conséquence une égalisation progressive des pressions partielles de part et d’autre de l’échangeur.
Le sang circulant dans les lamelles et les filaments, il est renouvelé de manière continue : le sang chargé en dioxygène et appauvri en dioxyde de carbone est évacué vers les artères branchiales efférentes et remplacé par du sang appauvri en dioxygène et chargé de dioxyde de carbone provenant des artères branchiales afférentes.
Ainsi la pression partielle de dioxygène est maintenue à un niveau bas et celle du dioxyde de carbone à un niveau élevé dans le milieu intérieur associé à l’échangeur.
La convection du milieu intérieur contribue à maintenir les gradients de pression partielle des gaz respiratoires au niveau de l’échangeur branchial, favorisant ainsi les transferts de gaz.
Les gradients moteurs de la diffusion dépendent des pressions partielles des gaz respiratoires dans le milieu intérieur mais également dans le milieu extérieur.
Comment sont-elles contrôlées ?
Une circulation d’eau unidirectionnelle
Des mouvements ventilatoires dus aux musculatures buccale et operculaire
Copyright 2019 – Perrine Giunchi ; Johanna Ribes
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.

Écouter/Voir la vidéo
au format .mp4
au format .webm
Au repos, le Vairon présente des mouvements coordonnés d’ouverture et de fermeture de la bouche d’une part, d’écartement et de rapprochement des opercules d’autre part.
Schématiquement dans une première phase, la bouche est ouverte et les ouïes sont fermées alors que les opercules sont écartés du corps. Dans une seconde phase, la bouche est fermée et les ouïes sont ouvertes alors que les opercules sont rapprochés du corps.
Quelles sont les conséquences de ces mouvements ?
Copyright 2019 – Perrine Giunchi ; Johanna Ribes
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
Écouter/Voir la vidéo
au format .mp4
au format .webm
Le dépôt de bleu de méthylène devant la bouche du Vairon est suivi d’une expulsion d’eau colorée par les ouïes. Le bleu de méthylène circule donc de la cavité buccale à la cavité branchiale.
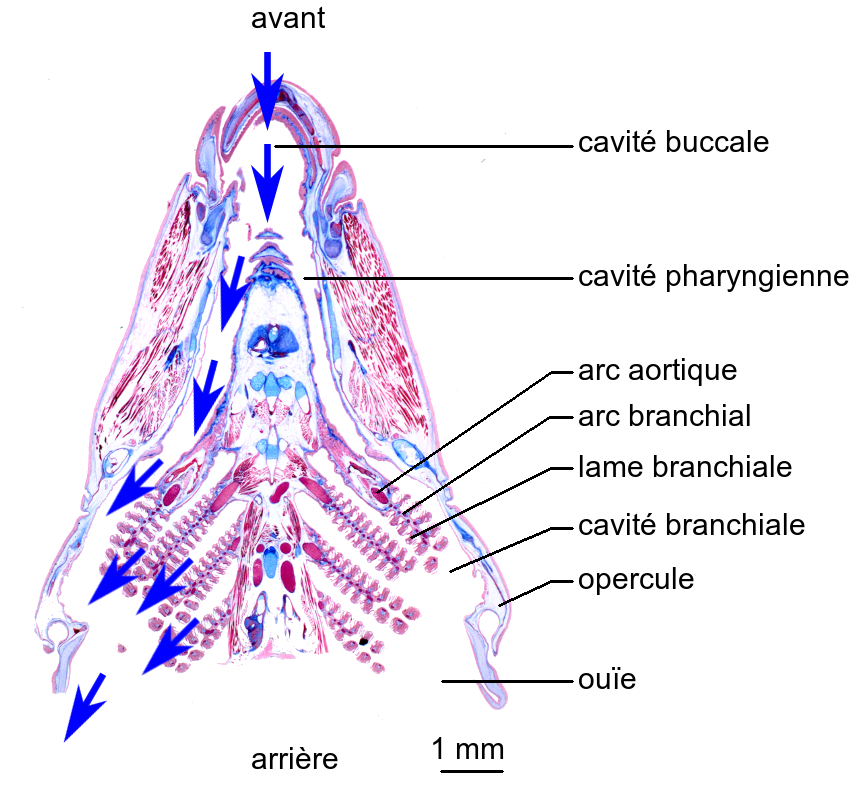
Copyright : 2019 – Pierre Hirt ; Perrine Giunchi ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
Le flux de bleu de méthylène révèle l’existence d’un courant d’eau dans la région antérieure du Vairon. L’eau entre dans la cavité buccale par la bouche, transite dans le pharynx puis pénètre dans les cavités branchiales à la faveur des fentes pharyngiennes et baigne les branchies avant de sortir par les ouïes.
La circulation de l’eau ainsi décrite est appelée ventilation. Réalisée en sens unique, la ventilation est qualifiée d’unidirectionnelle.
Comment l’eau est-elle mise en mouvement ?
Alors que la bouche est ouverte, le plancher buccal est abaissé du fait de l’action de sa musculature. Une dépression est générée dans la cavité buccale, provoquant une entrée d’eau.
Parallèlement, alors que les ouïes sont fermées, les opercules sont écartés en raison de l’action des muscles associés. Une dépression est également générée dans les cavités branchiales. Plus importante que la dépression de la cavité buccale, elle engendre une entrée d’eau depuis la cavité buccale.
Cette phase est dite d’expansion.
Inversement, lorsque la bouche est fermée, le plancher buccal est relevé. Une surpression est développée dans la cavité buccale, chassant l’eau dans le pharynx.
Simultanément, les ouïes sont ouvertes et les opercules rapprochés. Une surpression est également générée dans les cavités branchiales. Inférieure à celle de la cavité buccale, elle ne s’oppose pas au passage de l’eau à travers les fentes branchiales, mais assure sont évacuation par les ouïes.
Cette phase est dite de compression.
Ainsi un courant d’eau continu renouvelle le milieu extérieur au contact de l’échangeur branchial des Téléostéens, assurant le maintien des pressions partielles des gaz respiratoires.
Le caractère unidirectionnel de la ventilation est interprété comme économe en énergie au regard de la viscosité et de la densité élevée de l’eau.
Chez le Thon et le Maquereau par exemple, la ventilation est également assurée par la nage, bouche ouverte.
Qu’en est-il chez les autres Vertébrés ?
Des mouvements ventilatoires dus aux musculatures buccale et branchiale
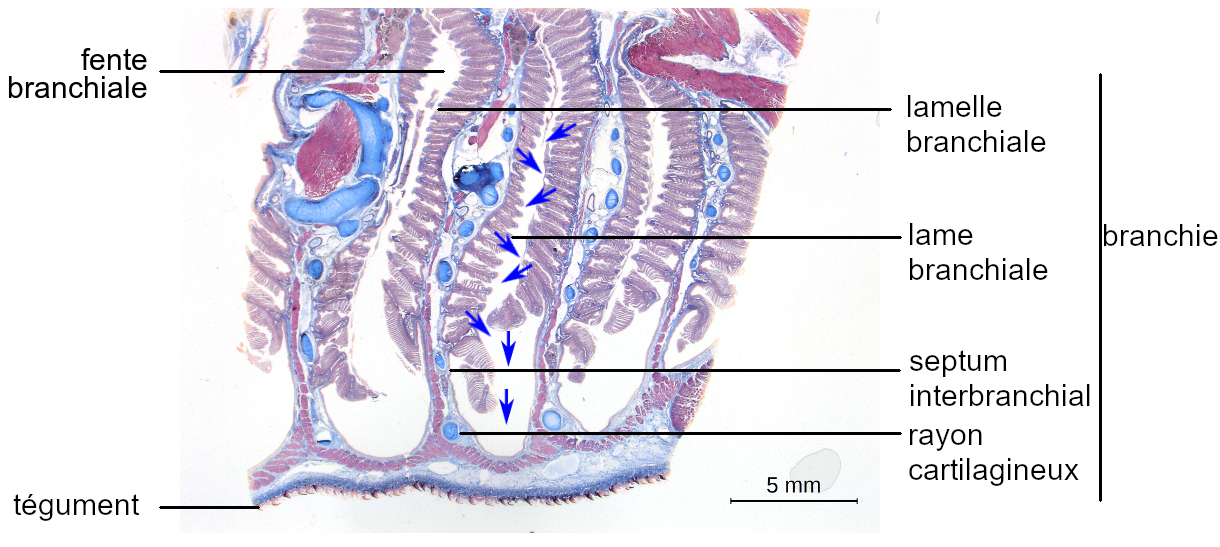
Copyright : 2019 – Pierre Hirt ; Perrine Giunchi
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
Chez les Sélaciens de la même manière, un courant d’eau unidirectionnel renouvelle l’eau de la cavité branchiale. Les espèces benthiques possèdent des orifices dorsaux, les spiracles, permettant l’entrée d’eau dans le pharynx, de même que la bouche.
Lorsque la bouche et les spiracles sont ouverts, les fentes branchiales sont fermées par l’extrémité des septums jouant le rôle de clapets. La cavité pharyngienne est dilatée par l’action des muscles associés.
Une dépression est générée, provoquant l’entrée d’eau dans le pharynx. Les clapets obturant les fentes branchiales sont écartés, et sont à l’origine d’une dépression plus importante dans la cavité branchiale, autorisant l’entrée d’eau depuis le pharynx.
Inversement, lorsque la bouche et les spiracles sont fermés, les fentes branchiales sont ouvertes. La cavité pharyngienne est comprimée par l’action des muscles associés et l’eau est chassée à travers les fentes pharyngiennes. La musculature des septums provoque de même la compression des chambres branchiales et l’expulsion de l’eau par les fentes branchiales.
La pression plus élevée dans le pharynx que dans les chambres branchiales garantit un écoulement d’eau permanent au contact des lamelles branchiales.
Certains Sélaciens nagent également bouche ouverte, leur ventilation est alors liée à leur progression dans l’eau, due à l’action de la musculature somatique.
De manière générale, la ventilation des branchies internes des Vertébrés est unidirectionnelle et due à l’action de la musculature de la région antérieure ou des branchies elles-mêmes. Les Lissamphibiens ne font pas exception.
Qu’en est-il des branchies externes ?
Une ventilation sans direction privilégiée ou bidirectionnelle
La ventilation des branchies externes des larves de Lissamphibiens est simplement réalisée par leur agitation dans l’eau. Elle est due aux contractions de la musculature associée aux arcs branchiaux.
Une ciliature superficielle est parfois présente, ses battements sont responsables d’un déplacement local de l’eau.
La ventilation ne présente pas dans ce cas de direction privilégiée.
Les Lamproies adultes constituent un cas original dans la mesure où elles possèdent un appareil branchial interne représenté par des poches branchiales. Parasites, elles sont fixées à leur hôte par la bouche, qui ne peut en conséquence être l’orifice par lequel l’eau pénètre au cours de la ventilation.
L’eau entre et sort des poches branchiales par des spiracles, orifices latéraux. Elle est mise en mouvement par l’action de leur musculature notamment.
La ventilation est dans ce cas bidirectionnelle.
Quelles que soient ses modalités, la ventilation permet le maintien des pressions partielles des gaz respiratoires à un niveau constant dans le milieu extérieur, et contribue ainsi au maintien des gradients de pression partielle de part et d’autre de l’échangeur.
Comment sont orientées les convections des milieux extérieur et intérieur l’une par rapport à l’autre ?
Des convections des milieux intérieur et extérieur à contre-courant
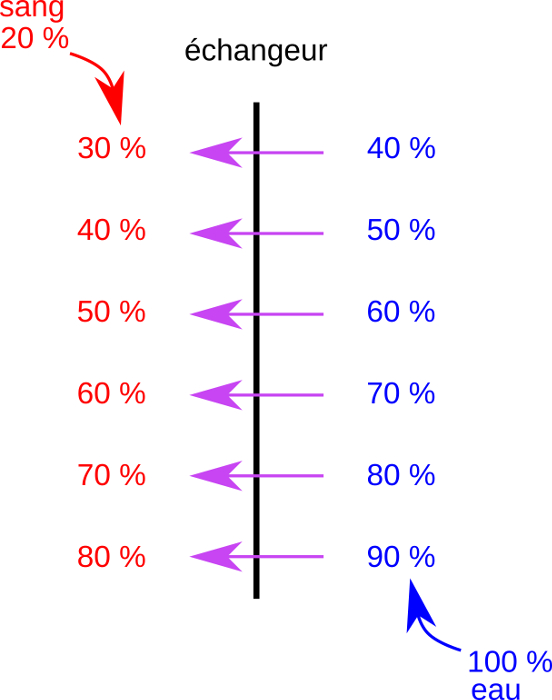
Copyright : 2019 – Ismaël Bamana ; Sandrine Heusser
Cette illustration est mise à disposition selon les termes de la Licence Creative Commons Attribution – Pas d’Utilisation Commerciale – Partage dans les mêmes conditions 4.0 International.
Chez les Téléotéens, le sang des lamelles branchiales et l’eau de la cavité branchiale circulent en sens opposés.
Cette disposition est dite à contre-courant.
Dans ces conditions, le sang se trouve toujours en présence d’une eau dans laquelle la pression partielle du dioxygène est supérieure. En conséquence le dioxygène diffuse du milieu extérieur au milieu intérieur sur toute la longueur de l’échangeur et le taux d’extraction de ce gaz est élevé.
La circulation à contre-courant favorise l’extraction du dioxygène car elle est à l’origine d’un gradient de pression partielle constant tout au long de l’échangeur.
Elle est interprétée comme une adaptation à la quantité de dioxygène relativement faible dans l’eau.
Finalement, les Vertébrés possédant un appareil respiratoire branchial réalisent de manière générale une ventilation unidirectionnelle. Convection renouvelant le milieu extérieur au contact de l’échangeur, elle favorise la diffusion des gaz respiratoires au même titre que la convection du milieu intérieur.
En milieu aquatique, les appareils branchiaux des Vertébrés sont fréquemment le siège d’échanges ioniques.